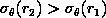From the point simulations within spheres and shells, for
different
(FWHM) is given in figure 5.4, 5.5,
5.6 and 5.7 for
and r ranging from 15 to 90
mm. As would be expected, it shows that the translation errors are
independent of configuration dimension for the range of homology errors
that may be expected clinically because the translation transformations
necessary for registration are simply calculated as the difference between
centroid positions (refer to chapter on Procrustes analysis). The results
given are for the x dimension, but it sufficiently describes the behavior
in all three dimensions through symmetry arguments because the simulations
were performed within spherical geometries.

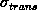
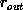

The same behavior is obtained from simulations within the
shells. It is of interest to point out that the results from
configurations within shells or spheres are essentially the same. In
general, the delineation of the data's dependence on radius is simply
less scattered for the simulations within shells. This may be seen
for in
figures 5.9, 5.10, 5.11 and 5.12 for both
spheres and shells. The curve gives a
Simplex
![]() fit to
fit to (where k is a constant
that is proportional to
and inversely proportional to
plus a constant). The bottom plot in figure 5.8 give
the values for the constant k resulting from the fit. The change in rotation
error with radius for spheres and shells is shown to be the same from
the correspondence of the k values resulting from the fits.
Although the development of an exact
functional dependence is not the purpose here, this
correspondence clearly demonstrates the decrease of
with increase of r. The deviation of the data points to the curve may
be seen to be generally greater for spheres than for shells because
the shells constrain the radial dependence more tightly than the
spheres. This corresponds to the standard deviation of the difference between
the fits and the simulation data being larger for the spheres than for
the shells. This may be seen in the upper plot of figure 5.8.
For small perturbations where
mm, both
and
are fairly independent of point
set configuration. This has previously been shown theoretically in
terms of the Procrustes statistic [Sib79].
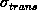
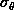
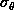
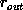
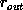
The dependence of the errors on the number of points used is well
established. Studies show that they decrease by
[NCH+92][HHH+92]. It is clear from the figures that the uncertainty
in the rotation error depends very greatly on the radial dispersion of the
points from their centroid.
is greater for smaller
radii. This is due to the larger lever introduced by the same homology
error when the points are nearer the centroid (see figure 5.13).
The independence of
on the boundary used to constrain the
radial dependence, either shell or sphere, is of interest. This suggests
that there is no significant advantage to limiting the selection of point
pairs to shell-like configurations, corresponding to points selected on
the surface of the brain (refer to constants, k, in the bottom part of
figure 5.8).
This is contrary to what has been suggested
elsewhere [HHH+92]. The results from points simulations
within spheres and shells indicate that the homology error does affect
and
dramatically. Changes in both
and
are most dependent on the range of
the expected variation of
as well as by the differences
and possible range of r and m.